

POTENSI TRICHODERMA MENINGKATKAN KETAHANAN TANAMAN TERHADAP KEKERINGAN
Pendahuluan
Perubahan iklim menjadi salah satu faktor pembatas dalam upaya peningkatan produksi baik tanaman pangan maupun hortikultura. Salah satu dampak perubahan iklim tersebut yang saat ini terjadi adalah fenomena pemanasan Suhu Muka Laut (SML) di atas kondisi normalnya atau dikenal dengan El Nino. El Nino meningkatkan potensi pertumbuhan awan di Samudra Pasifik Tengah dan mengurangi curah hujan di Wilayah Indonesia. El Nino ini memicu terjadinya kondisi kekeringan di Wilayah Indonesia secara umum. Kekeringan merupakan salah satu faktor cekaman abiotik yang dapat menghambat pertumbuhan tanaman serta menurunkan hasil produksi padi. Kondisi kekeringan pada fase pembungaan akan berdampak terhadap menurunnya jumlah dan ukuran bulir padi. Cekaman kekeringan juga dapat menghambat rantai distribusi karbohidrat dari daun ke bulir sehingga berdampak pada menurunnya jumlah dan bobot bulir. Kekeringan juga mempengaruhi ketersediaan unsur hara dan transport unsur hara dari akar ke pucuk (Hudan Schmidhalter, 2005).
Apabila cekaman kekeringan berkepanjangan maka tanaman akan mati. Kekurangan air akan mengganggu aktifitas fisiologis maupun morfologis, sehingga mengakibatkan terhentinya pertumbuhan. Defisiensi air yang terus menerus akan menyebabkan perubahan irreversibel (tidak dapat balik) dan pada gilirannya tanaman akan mati. Meskipun demikian, toleransi tanaman terhadap cekaman kekeringan berbeda-beda dan mekanisme yang terlibat dalam ketahanan tanaman juga berbeda. Tanaman memiliki mekanisme yang potensial untuk bertahan terhadap tekanan kerusakan induksi oksidatif melalui sistem pertahanan antioksidan (Hashem et.al. 2016). Selain itu, kemampuan tanaman dalam mentolerir cekaman abiotik juga bergantung kepada asosiasi tanaman dengan mikroba seperti mikoriza, mikroba perakaran, mikroba pemacu pertumbuhan rizobacteria, mikroba endofit juga berperan penting dalam proses fisiologi tumbuhan (Ali et.al. 2014). Di antara beberapa jenis mikroba tersebut Trichoderma spp. dilaporkan memiliki potensi untuk meningkatkan toleransi tanaman terhadap cekaman biotik dan abiotik, termasuk kekeringan dan salinitas (Ahmad et.al. 2015).
Trichoderma spp. Meningkatkan Ketahanan Tanaman Terhadap Kekeringan
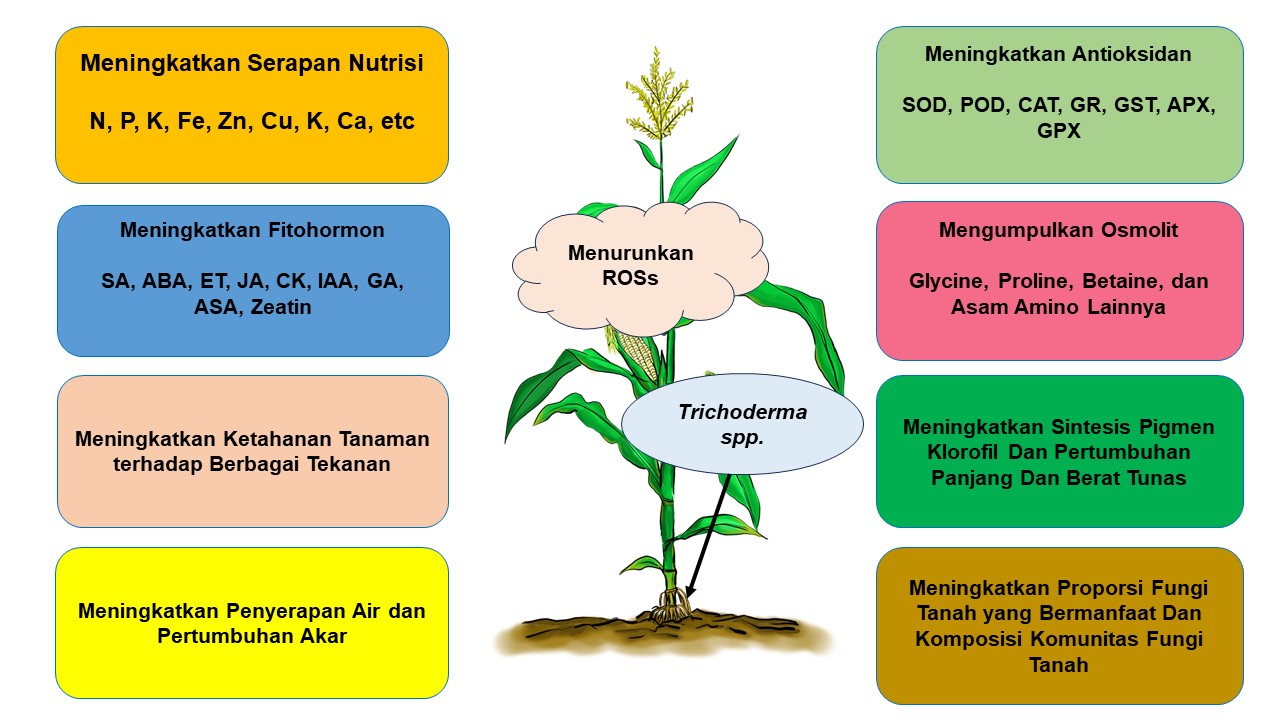
Trichoderma spp. merupakan salah satu cendawan antagonis yang telah banyak digunakan dalam berbagai praktek budidaya tanaman, baik sebagai agens biokontrol maupun sebagai pemacu pertumbuhan tanaman. Trichoderma spp. dipercaya sebagai agen biokontrol yang unggul karena karakternya yang unik, seperti kemampuan berkembang biak yang cepat, penyebaran yang tinggi dan kemudahan dalam isolasi dan perbanyakan. Trichoderma spp. umumnya hidup bebas dan sangat interaktif di lingkungan perakaran, tanah dan daun tanaman (Harman and Kubicek, 2020). Beberapa hasil penelitian menunjukkan bahwa kolonisasi Trichoderma di perakaran tanaman dapt meningkatkan pertumbuhan akar tanaman di bawah cekaman kekeringan. Trichoderma juga memiliki peran sebagai biokontrol terhadap patogen tanaman dan saat ini populer sebagai pemacu pertumbuhan tanaman (Chepsergon et al., 2014).
Trichoderma membentuk hubungan simbiosis dengan tanaman inang pada fase ketiga dalam proses perkecambahan biji hingga tanaman memasuki fase penuaan (Hermosa et al., 2012). Trichoderma membantu tanaman dalam menghindari cekaman kekeringan melalui adaptasi morfologi. Kolonisasi akar tanaman oleh cendawan Trichoderma harzianum dapat memodulasi hormon endogen tanaman, enzim ketahanan, antioksidan dan senyawa fenol yang dapat meningkatkan kemampuan ketahanan dibawah cekaman kekeringan (Chen et al., 2013). Trichoderma spp. memberikan efek langsung pada tanaman, diantaranya peningkatan laju pertumbuhan, penyerapan nutrisi, meningkatkan persentasi perkecambahan dan ketahanan tanaman terhadap cekaman biotik serta abiotik (Shoresh et al., 2010). Gusain et. al. (2014) melaporkan bahwa kolonisasi Trichoderma harzianum T35 pada perakaran padi dapat meningkatkan toleransi tanaman padi terhadap kekeringan. T. harzianum memacu aktivitas enzim antioksidan, superoksida dismutase (SOD), katalase (CAT), ascorbate peroksidase (APX) dan dengan demikian mencegah kerusakan oksidatif pada tanaman padi melalui eliminasi reactive oxygen species (ROS).
Hasil yang sama dilaporkan oleh Sutrisno et.al (2022) bahwa perlakuan Trichoderma harzianum pada berbagai konsetrasi secara signifikan dapat meningkatkan pertumbuhan dan hasil tanaman kedelai dan terdapat interaksi antara nyata antara keduannya. Perlakuan dosis 40 g Trichoderma dan cekaman kekeringan 60% menghasilkan pertumbuhan tanaman kedelai paling tinggi (72,92 cm) dibandingkan dengan tanpa perlakuan Trichoderma harzianum dengan tinggi tanaman 37 cm. Demikian pula pada variabel panjang akar tanaman yang diberi perlakuan Trichoderma harzianum menghasilkan panjang akar 8.20 cm dibandingkan dengan tanpa perlakuan dengan panjang akar 4.59 cm. Hasil tersebut menunjukkan bahwa aplikasi Trichoderma mampu menginduksi hormon endogen dalam tanaman kedelai untuk mendorong pemanjangan akar pada kondisi cekaman kekeringan (Chen et al., 2013). Kolonisasi Trichoderma pada akar tanaman akan meningkatkan ketahanan perakaran tanaman, memacu pertumbuhan dan perkembangan tanaman serta ketahanan terhadap cekaman kekeringan (Bae et al., 2009). Trichoderma juga mampu memfasilitasi kolonisasi akar, meningkatkan koordinasi mekanisme pertahanan tanaman dan meningkatkan laju fotosintesisi pada daun (Vargas et al., 2009).
Hasil penelitian tersebut juga menunjukkan bahwa perlakuan Trichoderma harzianum pada tanaman kedelai signifikan meningkatkan kandungan prolin yang merupakan osmoprotectant pada tanaman untuk meningkatkan ketahanan tanaman terhadap cekaman kekeringan. Trichoderma harzianum mampu mendorong tanaman untuk menghasilkan senyawa prolin lebih tinggi dibandingkan tanaman tanpa aplikasi Trichoderma harzianum. Kandungan prolin yang tinggi pada tanaman dengan cekaman kekeringan berkaitan erat dengan fungsi prolin sebagai senyawa osmoprotectant yang dihasilkan oleh tanaman kedelai sebagai bentuk ketahanan akibat cekaman kekeringan. Akumulasi senyawa prolin dipergunakan sebagai indikasi tanaman memiliki sifat toleran terhadap cekaman kekeringan yang dapat menurunkan tekanan air sel sehingga tanaman menyerap air dan hara lebih banyak di dalam tanah (Chen et al., 2013).
Mekanisme toleransi tanaman terhadap cekaman kekeringan antara lain tanaman akan mengakumulasi senyawa yang berfungsi untuk melindungi sel dari kerusakan dengan cara mengatur potensial osmotik sel. Senyawa yang berperan dalam penyesuaian osmotik sel adalah kandungan gula total dan senyawa prolin. Senyawa-senyawa tersebut dapat menurunkan potensial air di dalam sel dengan tanpa membatasi fungsi dari pada enzim dan menjaga tekanan turgor sel (Nazirah, 2018). Indikator tanaman dikatakan toleran terhadap cekaman kekeringan ditandai dengan adanya peningkatan jumlah kandungan prolin. Fungsi dari senyawa prolin adalah sebagai penyimpan unsur nitrogen, osmoregulator, dan protektor enzim tertentu. Senyawa prolin juga berfungsi sebagai penjaga turgor sel dan pertumbuhan akar pada saat kondisi potensial osmotik air rendah (Nurmalasari, 2018).
Berbagai hasil penelitian lainya juga menyebutkan bahwa inokulasi Trichoderma spp. pada tanaman dapat meningkatkan pertumbuhan akar, yang akan berdampak pada peningkatan penyerapan air dan unsur hara oleh akar, memperbaiki kecukupan air pada berbagai organ tanaman (Rawal et.al. 2022), dan melalui peningkatan konduksi stomata meningkatkan efisiensi tanaman dalam konsumsi air dan toleransi terhadap cekaman kekeringan (Scudeletti et.al. 2021). Inokulasi Trichoderma spp. pada tanaman padi dan jagung dengan metode priming benih (perlakuan pada benih) dilaporkan dapat meningkatkan pertumbuhan akar, meningkatkan efisiensi penyerapan air, meningkatkan ketahanan tanaman terhadap cekaman kekeringan (Scudeletti et.al. 2021). Berbagai spesies Trichoderma seperti T. viridie, T. asperelloides, T. virnes dan T. harzianum dilaporkan meningkatkan kualitas fotosintesis pada tanaman (Rawal et.al. 2022), meningkatkan kualitas dan kandungan pigmen fotosintesis, meningkatkan ketahanan tanaman, pertumbuhan berbagai organ tanaman, meningkatkan biomassa produksi tanaman pada cekaman kekeringan (Contreras-Cornejo, et.al. 2009). Mishra et.al. (2020) melaporkan bahwa aplikaksi Trichoderma spp. Pada tanaman padi meningkatkan akumulasi klorofil, pertumbuhan akar, peningkatan jumlah anakan dan meningkatkan hasil pada kondisi cekaman kekeringan.
Berbagai hasil penelitian di atas menunjukkan bahwa Trichoderma spp. yang banyak dijumpai di semua lahan pertanian memiliki potensi untuk di eksplorasi dan dikembangkan sebagai agens biokontrol, pemacu pertumbuhan, dan meningkatkan ketahanan tanaman terhadap cekaman kekeringan.
Kontributor : Tuminem (POPT Ahli Madya) & Widia Herhayulika (POPT Ahli Muda)
Daftar Pustaka
Ahmad P, Hashem A, Abd_Allah E F, Alqarawi A A, John R, Egamberdieva D, Gucel S. 2015. Role of Trichoderma harzianum in mitigating NaCl stress in Indian mustard (Brassica juncea L.) through antioxidative defense system. Frontiers in Plant Science, 6, 868.
Ali S, Charles T C, Glick B R. 2014. Amelioration of high salinity stress damage by plant growth-promoting bacterial endophytes that contain ACC deaminase. Plant Physiology & Biochemistry, 80, 160–167.
Bae, H., Sicher, R. C., Kim, M. S., Kim, S. H., Strem, M. D. et al. (2009). The beneficial endophyteTrichoderma hamatum isolate DIS 219b promotes growth and delays the onset of the drought response in Theobroma cacao. Journal of Experimental Botany, 60(11), 3279–3295
Contreras-Cornejo, H. A., Macías-Rodríguez, L., Cortés-Penagos, C., López-Bucio, J. (2009). Trichoderma virens, a plant beneficial fungus, enhances biomass production and promotes lateral root growth through an auxin-dependent mechanism in Arabidopsis. Plant Physiology, 149(3), 1579–1592.
Chepsergon, J., Mwamburi, L., & Kassim, M. K. (2014). Mechanism of Drought Tolerance in Plants Using. International Journal of Science and Research (IJSR), 3(11), 1592–1595.
Chen, Q., Tao, S., Bi, X., Xu, X., Wang, L., & Li, X. (2013). Research progress in physiological and molecular biology mechanism of drought resistance in rice. American Journal of Molecular Biology, 03(02), 102–107.
Hashem A, Abd_Allah E F, Alqarawi A A, Al-Huqail A A, Wirth S, Egamberdieva D. 2016. The Interaction between arbuscular mycorrhizal fungi and endophytic bacteria enhances plant growth of Acacia gerrardii under salt stress. Frontiers in Microbiology, 7, 1089.
Hermosa, R., Viterbo, A., Chet, I., & Monte, E. (2012). Plant-beneficial effects of Trichoderma and of its genes. Microbiology, 158(1), 17–25.
Mishra, D., Rajput, R. S., Zaidi, N. W., Singh, H. (2020). Sheath blight and drought stress management in rice (Oryza sativa) through Trichoderma spp. Indian Phytopathology, 73(1), 71–77.
Nazirah, L. 2018. Teknologi Budidaya Padi Toleran Kekeringan. CV. Sefa Bumi Persada, Aceh.
Nurmalasari, IR. 2018. Kandungan asam amino prolin dua varietas padi hitam pada kondisi cekaman kekeringan.Agrotech Science Journal. 4: 29–44.
Rawal, R., Scheerens, J. C., Fenstemaker, S. M., Francis, D. M., Miller, S. A. et al. (2022). NovelTrichoderma isolates alleviate water deficit stress in susceptible tomato genotypes. Frontiers in Plant Science, 13, 869090.
Shoresh, M., Harman, G. E., & Mastouri, F. (2010). Induced systemic resistance and plant responses to fungal biocontrol agents. Annual Review of Phytopathology, 48, 21– 43.
Vargas, W. A., Mandawe, J. C., & Kenerley, C. M. (2009). Plantderived sucrose is a key element in the symbiotic association between Trichoderma virens and maize plants. Plant Physiology, 151(2), 792–808